MY RESEARCH particularly focuses on estimating key life history parameters required in population dynamic growth models of marine top predators to elucidate:
i) the importance of biotic and abiotic factors that influence growth, condition, and survival.
ii) the population dynamics-level responses to environmental change.
iii) the ecological implications of life history parameters into species conservation status.
As such, my research employs a large-spectrum of analytical methods and innovative technologies under a multidisciplinary frame including bio-chronology, microsatellites DNA markers, natural tags (microchemistry), distribution, abundance and acoustic data. Specifically, my research is broadly motivated to explore and understand different skeletal hard structures (dolphin tooth, fin spines and otoliths) used for aging beyond their chronological properties.
The following briefly describe the major projects being undertaken during my research career.
i) the importance of biotic and abiotic factors that influence growth, condition, and survival.
ii) the population dynamics-level responses to environmental change.
iii) the ecological implications of life history parameters into species conservation status.
As such, my research employs a large-spectrum of analytical methods and innovative technologies under a multidisciplinary frame including bio-chronology, microsatellites DNA markers, natural tags (microchemistry), distribution, abundance and acoustic data. Specifically, my research is broadly motivated to explore and understand different skeletal hard structures (dolphin tooth, fin spines and otoliths) used for aging beyond their chronological properties.
The following briefly describe the major projects being undertaken during my research career.
Life history and growth of marine top predators
Age determination of small odontocetes
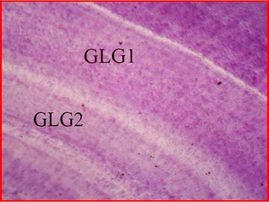
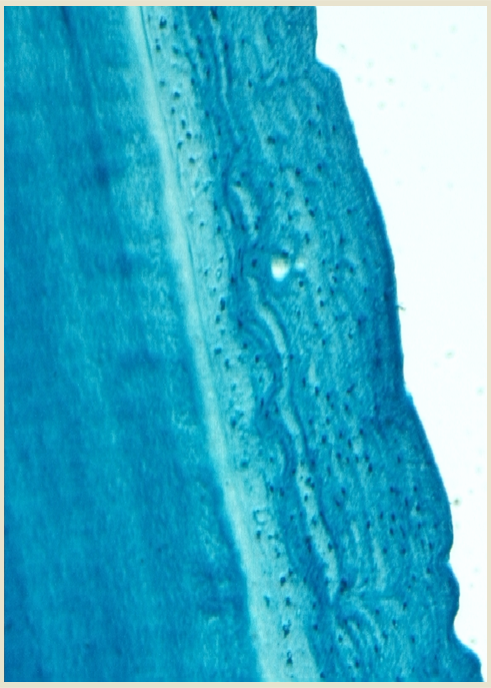
Age estimation in odontocetes is based on counts of Growth Layers Groups (GLGs) deposited in recording structures such as teeth (Perrin & Myrick 1980), similar in concept to reading growth rings in trees.
A GLG generally consists of a bipartite structure that includes alternating wide stainable layer follow by a narrow non-stainable layer in decalcified and stained sections. The observed contrast is due to differences in content and distribution of the inorganic matrix (hydroxyapatite) resulting in differences in optical density and stainability, with the more transparent bands containing more mineral compounds.
These GLSs deposited parallel to the formative surface of a tissue occur with cyclical and predictable repetition and hence they are assumed to be laid down annually as a result of seasonal changes in the growth rate of the tooth, which are correlated to seasonal changes in the animal´s growth rate.
A GLG generally consists of a bipartite structure that includes alternating wide stainable layer follow by a narrow non-stainable layer in decalcified and stained sections. The observed contrast is due to differences in content and distribution of the inorganic matrix (hydroxyapatite) resulting in differences in optical density and stainability, with the more transparent bands containing more mineral compounds.
These GLSs deposited parallel to the formative surface of a tissue occur with cyclical and predictable repetition and hence they are assumed to be laid down annually as a result of seasonal changes in the growth rate of the tooth, which are correlated to seasonal changes in the animal´s growth rate.
Comparison of two histological techniques for age determination in small cetaceans
Decalcified and stained thin tooth sections is the most widely method used for age determination in odontocetes, particularly for those species that have small teeth, allowing accurate age estimates (Hohn and Fernández 1999). Tooth sections are generally obtained using a freezing microtome or cryostat, but little information has been published on the use of alternative methods such as the standard paraffin microtome.
This project aims to investigate whether the paraffin method can be a viable alternative to the traditional cryostat for preparing dolphin teeth and obtaining reliable age readings.




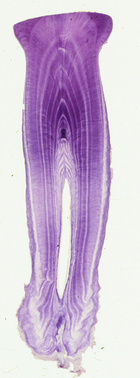
Teeth of several small odontocete species including Harbour porpoises (Phocoena phocoena) or common dolphin (Delphinus delphi), were processed using both techniques, testing different section thicknesses (8, 16, 24μm) and staining methods (Mayer’s, Ehlrich’s haematoxylin, Toluidine blue and Giemsa). Comparative analysis within and between techniques were carried out to investigate whether these factors contributed significantly to the quality ("readability") of tooth sections, and the age interpretation. Differences between species were also tested to evaluate which technique would generally be working better for preparing small dolphin tooth ensuring a reliable estimated ages.
Results of this study show that particular effects of staining methods and/or sections thicknesses should be considered within each technique, however Mayer’s haematoxylin stained sections of 8μm thickness seems to provide the best contrast of the GLGs with a good age agreement for both techniques, with higher percentage of preparations classified as `good quality´ for the paraffin method. The major contribution of this work is that the paraffin method facilitates the cutting of thinner sections, increasing the chances of obtaining a tooth section closer to the pulp cavity´s center, which is particularly beneficial when the availability of tooth samples is constrained. In addition, this technique is accessible to most researchers,( i.e., it requires less specialist lab technician skills than the cryostat technique).
Hence, it is concluded that the Paraffin technique represents a viable and cost-effective alternative to the traditional cryostat for preparing dolphin teeth, and especially successful for small odontocete species such as porpoises. More details in Luque (PhD 2008), Luque et al., 2009. Marine Mammal Science 25(4),902-919.
Mineralization anomalies in teeth of small cetaceans: do they provide a record of key life history events and/or environmental stresses?
Annual GLGs very often can contain certain alterations (or disturbances) in the normal regular deposition of dental tissues. In odontocetes and sirenians, such tooth-tissue disturbances are termed mineralization anomalies. Five types of mineralization anomalies have been identified: accessory and marker lines are discrete regular layers noticeably different in appearance (e.g. staining light or dark) within a GLG; dentinal resorption and cemental disturbance refer to various degrees of mineralization interference-erosion and/or repair of existing regular laminated tissue; pulp stones are discrete nodules, often containing concentric rings of secondary dentine that generally appear in the region of the pulp cavity (Myrick 1988, Lockyer, 1993).

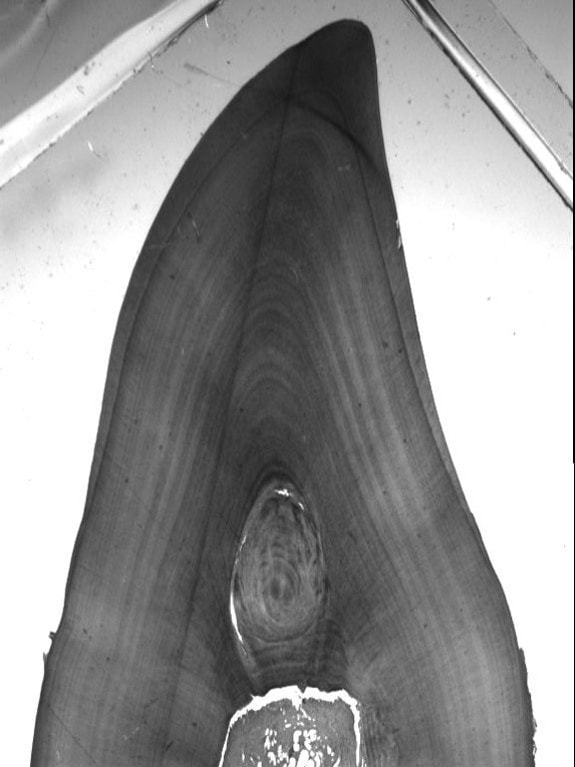
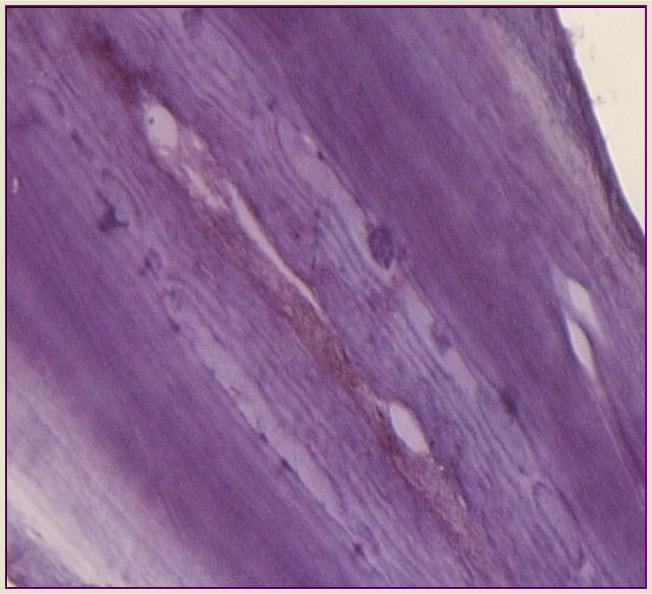
This project aims to examine the tooth ultra-structure of porpoises and common dolphins i) to describe the time course of accumulation of the distinct types of mineralization anomalies, and ii) to determine whether these anomalies might represent “time markers” of certain stressful conditions linked to life history events (i.e., age at attainment of sexual maturity) and/or bioaccumulation of anthropogenic organic pollutants (i.e., blubber POP concentrations).
Harbour porpoise
|
Common dolphin
|
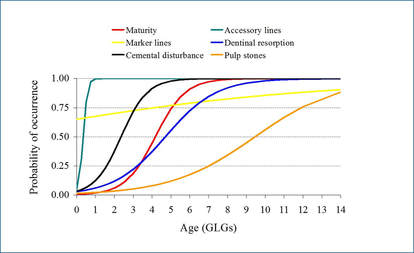
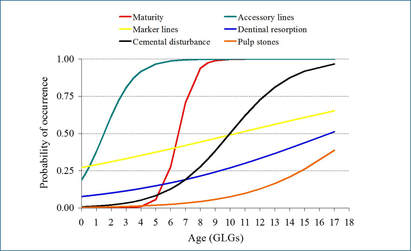
Fitted logistic curves for the incidence of anomalies in harbor porpoise (left) and common dolphins (right) in relation to age showing also the fitted maturity ogive (red curve)
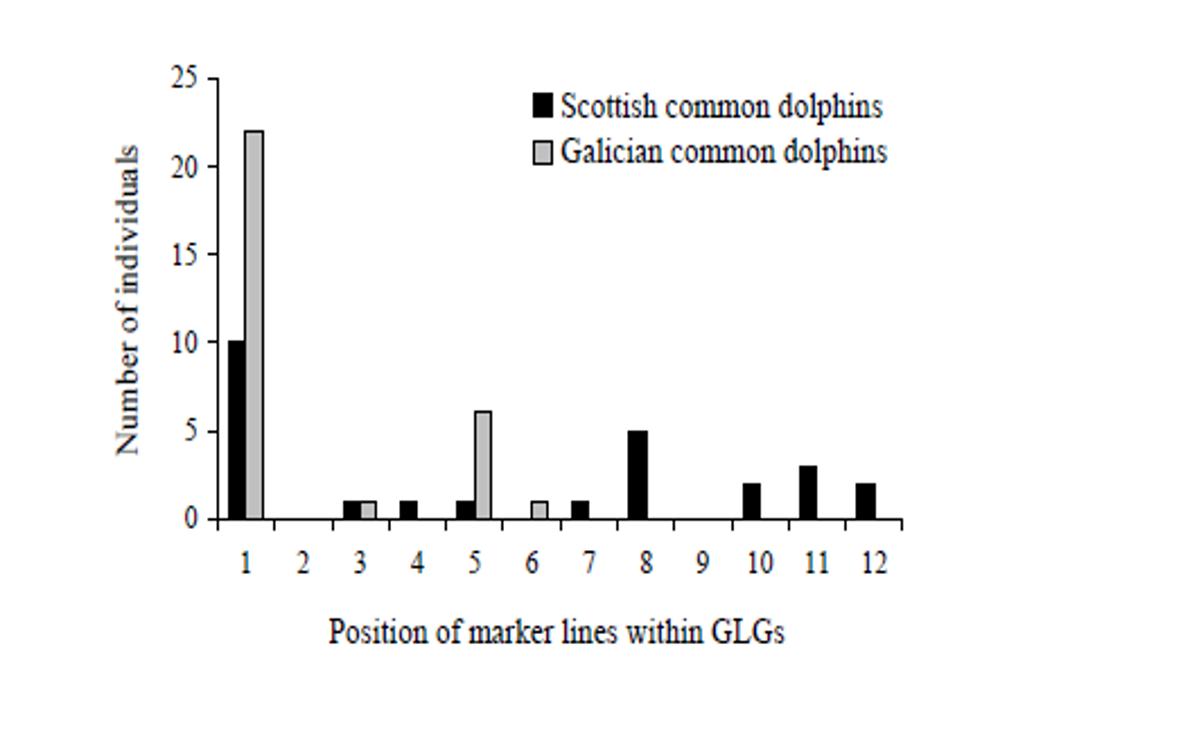
Results show that the presence of all tooth anomalies was overall high and increased with age, except for marker layers which show a less clear pattern in both species. The fact that the relationships between anomaly incidence and age tended to be logistic curves rather than straight lines implies that there are certain ages at which mineralization anomalies have a higher probability of occurring.
Fitted binary generalized linear and additive models (GLMs and GAMs, respectively) confirm that the three co-varying life history variables (i.e. age, length or maturity) are all significantly related to the presence of dentinal resorption and cemental disturbance in both species being `age´ the best predictor of the incidence of the two anomalies in porpoises while `length´ in common dolphins. The occurrence of dentinal resorption lagged behind maturation in common dolphins and porpoises while cemental disturbance tended to appear ahead maturation in porpoises. Hence, the time course of appearance of dentinal resorption and cemental disturbance was similar to the time course of maturation suggesting a link between anomaly occurrence and the age at which the animals become sexually mature in both species.
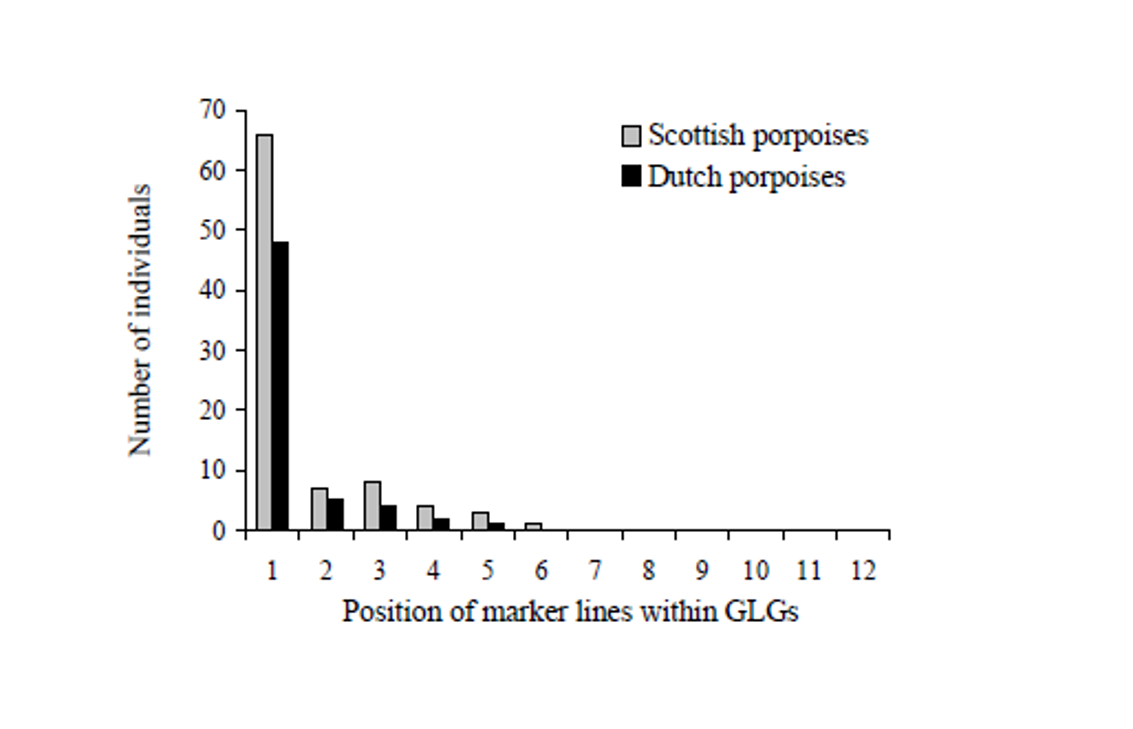
Moreover, there are two age ranges at which marker lines tended to appear more frequently, particularly high within the first GLG, close to the neonatal line, and then within 3rd-4th GLG in porpoises and 5th, 8th GLG in common dolphins, which coincide with the beginning of weaning and/or the age at sexual maturation, respectively (Learmonth et al., 2014, Murphy, 2004) suggesting an association with these two major life history events.
|
|
The major contribution of this work is that results provide evidence that certain tooth mineralization anomalies could be interpreted as time markers associated with life-history events, potentially representing a powerful tool for long-term monitoring and modelling.
Other concluding remarks:
Differences in the levels of incidence between sp. indicate that other factors (e.g., diet, temperature, etc.) could play an important role in their formation or simply varying degrees of susceptibility to the same causative factor. More details in Luque et la., 2009, 2013.
Other concluding remarks:
- [POPs] did not contribute to the incidence of anomalies neither in porpoises nor in commons dolphin and therefore they are not considered as useful indicators of stress associated with sexual maturity.
- The absence of sex differences suggests that parturition and lactation are not necessarily important stressors, or at least no more important than what males experience.
- The pattern of occurrence anomalies is similar for animals that are separated in time and space suggesting a systemic cause rather than a local one.
- The presence of mineralization anomalies is clearly a physiological process since all teeth examined per individual show the same stages of alteration.
Differences in the levels of incidence between sp. indicate that other factors (e.g., diet, temperature, etc.) could play an important role in their formation or simply varying degrees of susceptibility to the same causative factor. More details in Luque et la., 2009, 2013.
Age determination of Atlantic bluefin tuna
Ageing fish is one of the major activities of fisheries science due to individual fish growth is a fundamental biological process in population dynamics which are mostly age-based. The most common method to estimate growth, is the reading of hard parts (i.e. otoliths, fin spines, scales, or vertebra) to determine the annual growth increments of an individual fish providing direct age at length observations (Campana and Throrrold 2001).
In 2007, the BLUEAGE project framed within an agreement of scientific cooperation between the Fisheries and Ocean Canada (DFO) and the Spanish Oceanographic Institute (IEO), was signed to address the need to improve knowledge about Atlantic bluefin tuna life history in order to contribute to its management and ensure the sustainability of their populations. The main goal of BLUEAGE project is to validate bluefin tuna age interpretations using dorsal fin spines through bomb derived radiocarbon analysis that has already confirmed the accuracy of ages determined from otoliths (Neilson and Campana, 2008). Other aims of the project include i) review direct ageing methodology using fin spines, ii) to establish validated reference collections of paired hard structures (Rodriguez et al., 2014), and iii) to analyze temporal variability by modeling length at age from historic and current data series.
Direct ageing of Atlantic blue fin tuna (Thunnus thynnus) from the eastern North Atlantic and Mediterranean sea using dorsal fin spines
The main goal of this study is to expand the limited literature available on important methodological issues that affect direct age estimates using dorsal fin spines including:
- Definition of the spine in terms of its biometric relationships
- Definition sectioning location
- Quantification of spine nucleus vascularization
- Verification of the annual periodicity of annulus in spines to indirectly validate the spine-based age estimates
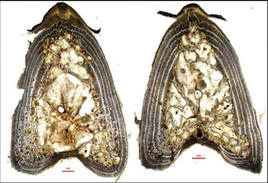
As the fish grow, the spine nucleus is reabsorbed replacing the earliest annuli by vascular tissue. This common process in large pelagic fish species is known as nucleus vascularization and is the greatest disadvantage of using this hard structure for ageing purposes, as it might result in significant age underestimation and growth overestimation (e.g., Drew et al., 2006; Kopf et al., 2010).
Nevertheless, very little has been published regarding the quantification of obscured annuli due to vascularization in most of these species including T. thynnus. The present study clearly shows that nucleus vascularization seriously affects specimens older than 6 years, with the disappearance of 40–50% of the presumed annuli by that age.
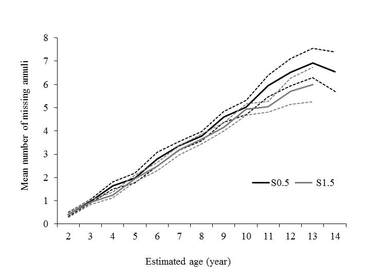
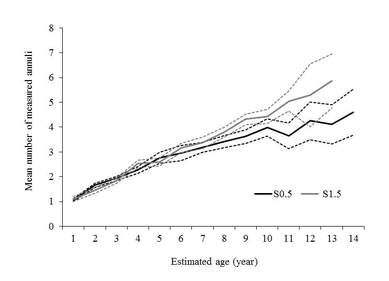
Exploring an alternate sectioning location s1.5, it certainly enhances the contrast of annulus providing a clear improvement regarding to the number of missing annulus due to vascularization that was significantly lower than the the traditional sectioning location S0.5 (Figure left). Besides, the number of measured annulus was significantly higher in the S1.5 than the S0.5 (Figure right), which represent an important contribution to the methodology of using this structure for ageing the full-length range of eastern T. thynnus. More details in Luque et al., 2014. Journal of fish biology 84, 1876-1903.
Bluefin tuna spine section showing the traditional S0.5 (left) and the alternate sectioning location S1.5 (right)
|
|
Age validation Research
Several assumptions must be met to effectively use hard parts for age and growth analysis: i) growth mark deposition must be deposited at a predictable time (e.g., daily or annually), and ii) must be readily identifiable. However, these assumptions are difficult to assess because consistency and clarity of growth mark deposition may change both within an individual (e.g., as fish become reproductively mature) and among populations due to environmental conditions. Hence, validating these assumptions is considered critical to use hard part structures for attaining information for age and growth.
DIRECT VALIDATION: Bomb radiocarbon analysis
Bomb radiocarbon derived from nuclear testing provides one of the best `direct´ age validation approaches available for long-lived fishes (Campana et al. 2008, Neilson and Campana 2008). The method uses anthropogenic carbon isotopes deposited within otoliths annular growth rings as a result of atmospheric nuclear weapons testing during the late 1950s as a dated mark. This method has successfully validated ages determined from bluefin tuna otoliths (Neilson and Campana, 2008), which resulted in a new growth curve for the western population (Restrepo et al., 2010). However, the lack of analogous validation studies for the eastern bluefin tuna population for which its current growth curve is based on estimates using fin spines (ICCAT, 2013) is the main motivation of BLUEAGE project. Thus, for the first time this study explores the potential value of bomb radiocarbon method for validating age interpretations in bluefin tuna spines available from 1984.
Specific objectives include i) to determine whether the radiocarbon content of spine annuli was consistent with expectations, and thus consistent with an accurate age interpretation, ii) to determine if there was evidence of contamination from the medium used to embed the spines. Results confirmed that fin spines contained radiocarbon at concentrations consistent with expectation. Moreover, spine radiocarbon chronology was consistent with an accurate age interpretation, but was also consistent with under-ageing.
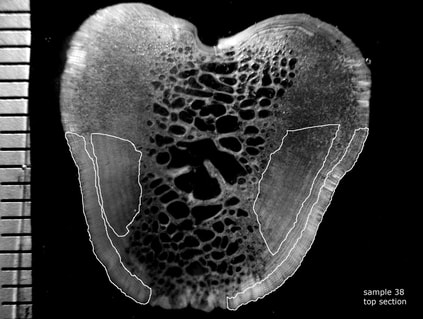
Fig. 1. Photomicrograph of a transverse section of a bluefin tuna fin spine showing presumed annuli (white dots) and regions isolated for radiocarbon assay (white polygons). Scale bar in 1-mm increments
|
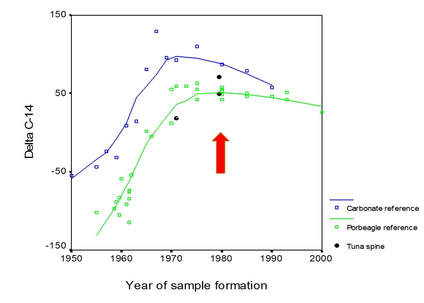
Fig. 2. Radiocarbon content of selected annuli in bluefin tuna fin spines (black symbols) compared to known-age radiocarbon reference chronologies based on carbonates (Arcto Norwegian cod otoliths; Kalish et al. 2001) and porbeagle shark vertebrae (Campana et al. 2002). Given that fin spines share the metabolic signature of vertebrae, the latter would be expected to more closely resemble the radiocarbon chronology of the spines.
|
INDIRECT VALIDATION: Edge type and Marginal Increment analysis
The main premise of these two indirect validation methods is that as fish age over an annual time-step, measurements of the outermost margin of the ageing structure .e., marginal increment ratio (MIR) or the proportion of opaque to translucent zones (i.e., edge analysis) will resemble a sinusoidal shape when plotted across months (Campana, 2001).
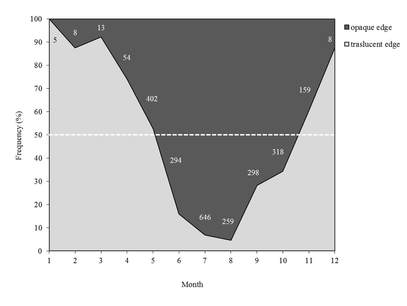
Fig.1. Categorization of edges by month for the T.thynnus (n = 2464). The horizontal dashed line indicates the 50% occurrence of edge type.
|
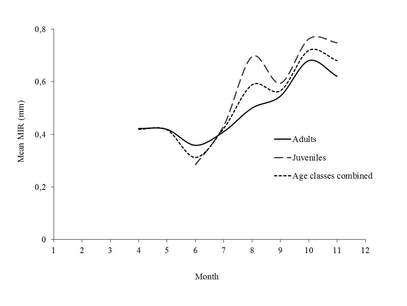
Fig. 2. Mean Marginal Increment ratio (MIR) by month for the T.thynnus for juveniles (ages 2–3; n = 423; dashed line), adults (ages > 3; n = 525; solid line) and all age classes combined (n = 948; dotted line).
|
The monthly categorization of bluefin tuna spine edge indicate that the highest frequency of translucent bands was low between May and October and then increased gradually during winter months until January and then decreased gradually (Fig.1 left). The frequency of opaque bands showed a reverse pattern of fluctuation, increasing during summer months, which suggests annual periodicity in the formation of the translucent bands. The monthly RMI showed two cycles in juveniles, but it clearly displayed an increasing trend that started in June to October regardless of age class (Fig.2 right) that coincided with the formation of opaque bands, which takes place mainly during summer months.
Results from edge type and marginal increment analyses in bluefin tuna indicate a yearly periodicity of annulus formation with the translucent bands (50% of occurrence) appearing from October to May.
Microchemical Analysis in fin spines
Trace Elements Analysis (TEA) of dorsal fin spines
Trace elements analyisis (TEA) that is commonly conducted on calcified structures (e.g., otolith, scales, fin rays, etc.,) is primarly used to track movements, habitat use, stock structure (e.g., Tzadik et al., 2017 and references herein). Otoliths have been the preferred bony structure to use in microchemical studies due to the fact that they are metabolically inert calcium carbonate structures. However, otolith removal requires sacrificing the fish, which is neither allowed for rare and/or endangered species, nor practical for commercially valuable fish species such as Atlantic bluefin tuna as it affects the appearance of the fish diminishing its market value. Here, we explore the suitability of dorsal fin spines of Atlantic bluefin tuna as a non-lethal and minimally invasive structure alternative to otoliths for trace elements analysis.
Trace elements are commonly analyzed by laser ablation-inductively coupled plasma-mass spectrometry (LA-ICP-MS) that produces an elemental fingerprint at a discrete time-point in the life of a fish, providing data on within-"hard" structure variability of that fingerprint. In general, little attention has been paid to the use of appropriate experimental designs in the scientific context of otolith studies. The presence of within-otolith variability has been acknowledged (e.g., Ruttenberg et al., 2005) and very recently incorporated into experimental designs aimed to assessing differences in elemental fingerprint of fishes from multiple sites on the presumed grounds (Di Franco et al., 2015). In this context, knowledge of within-fin spine variability is a key step crucial to assessing within fin-spine variability, and providing that within-fin spine measurements can be treated as ‘independent and identically distributed’ replicates.
Since the annuli on the fin spines show a concentric distribution, we use a hierarchical sampling design of spatial-temporal variation at four line scanning-ablation transects drawn from the nucleus to the edge to explore:
1) whether "multiple ablation spots within each seasonal annulus (i.e., translucent and opaque bands of each annulus) can be used as "independent" replicates for significance differences tests in the tracer concentrations between the opaque and translucent bands within/among annulus.
2) which are the main factors (i.e., band and/or year) contributing to variation in tuna fin spine microchemistry.
3) If so, to explore which of these tracers are contributing more importantly to such variation.
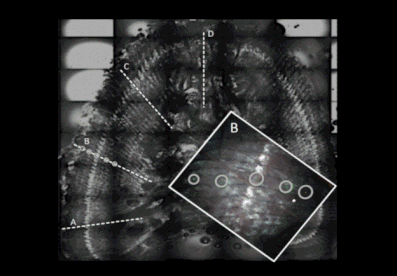
The photo (above) illustrate a cross-spine section showing the four line scanning ablation transects used onto each spine section. The zoom area shows that fin spines are ablated at multiple spots, equally spaced, ensuring that either the translucent and opaque bands were targeted along each ablation transect that are marked out prior to analysis within the laser setting program. The number of ablation spots in each spine section is constrained by the ablation crater diameter and the width of the translucent and opaque bands for each individual spine.
Trace elements are commonly analyzed by laser ablation-inductively coupled plasma-mass spectrometry (LA-ICP-MS) that produces an elemental fingerprint at a discrete time-point in the life of a fish, providing data on within-"hard" structure variability of that fingerprint. In general, little attention has been paid to the use of appropriate experimental designs in the scientific context of otolith studies. The presence of within-otolith variability has been acknowledged (e.g., Ruttenberg et al., 2005) and very recently incorporated into experimental designs aimed to assessing differences in elemental fingerprint of fishes from multiple sites on the presumed grounds (Di Franco et al., 2015). In this context, knowledge of within-fin spine variability is a key step crucial to assessing within fin-spine variability, and providing that within-fin spine measurements can be treated as ‘independent and identically distributed’ replicates.
Since the annuli on the fin spines show a concentric distribution, we use a hierarchical sampling design of spatial-temporal variation at four line scanning-ablation transects drawn from the nucleus to the edge to explore:
1) whether "multiple ablation spots within each seasonal annulus (i.e., translucent and opaque bands of each annulus) can be used as "independent" replicates for significance differences tests in the tracer concentrations between the opaque and translucent bands within/among annulus.
2) which are the main factors (i.e., band and/or year) contributing to variation in tuna fin spine microchemistry.
3) If so, to explore which of these tracers are contributing more importantly to such variation.
The photo (above) illustrate a cross-spine section showing the four line scanning ablation transects used onto each spine section. The zoom area shows that fin spines are ablated at multiple spots, equally spaced, ensuring that either the translucent and opaque bands were targeted along each ablation transect that are marked out prior to analysis within the laser setting program. The number of ablation spots in each spine section is constrained by the ablation crater diameter and the width of the translucent and opaque bands for each individual spine.
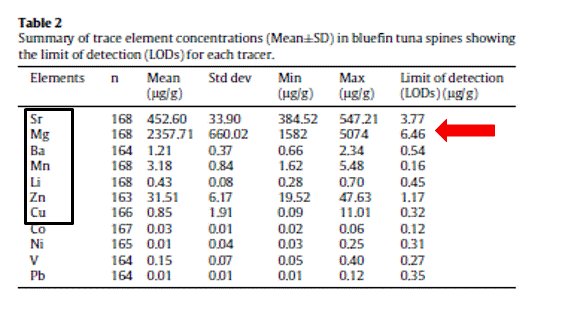
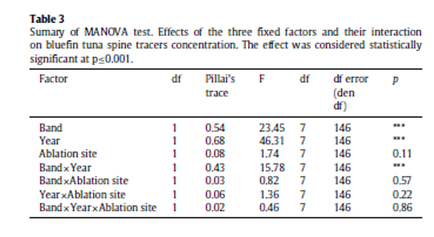
A total of seven out of eleven trace elements are consistently above the LODs in Atlantic bluefin tuna spines including Sr, Ba, Mg, Mn and Zn. (Table 2). Results of MANOVA indicate that elemental fingerprints measured in all multiple spots within seasonal annulus do not vary significantly among the four scanning-ablation transects, suggesting certain spatial and temporal stability of some tracer concentrations across the bipartite annular structure of the spine. As such, using multiple ablation spots as replicates, Pillai’s test show that the two fixed factors, i.e., band and year and their interaction have a significant effect in the overall spine elemental composition (p≤0.001) (Table 3).
|
|
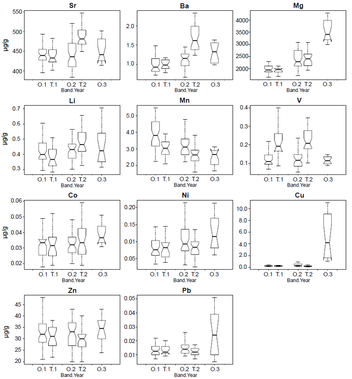
In conclusion, findings here provide an important insight regarding the metabolic stability of some tracer concentrations across the growth bands of the spine. Physiological reworking of the soft calcium phosphate (‘hidroxyapatite’) which constitutes the corpus calcareum of the spines is likely to be minimal, strengthening its use as a non-invasive alternative structure in elemental markers for this species. Using tracers in spine chemistry as a proxy of environmental conditions (e.g. temperature, salinity and elemental concentrations), and consequently for migration patterns studies, will depend on a more robust seawater chemistry characterization and further research on the elements incorporation mechanisms into the hard structure. Hence, it is crucially important to do more experimentation under controlled conditions to explore and test how tracers are bound within spine carbonate hydroxyapatite. Further research should also include older specimens where resorption has already taken place and the resorption effect on these trace elements.
Elemental concentrations of dorsal fin spines from Atlantic bluefin tuna (Thunnus thynnus) display by band (O=opaque, T =translucent) and year (i.e.annuli 1, 2 and 3). Whiskers range from 10th to 90th percentiles. Concentrations are given in (lg/g).
Genetic Population Structure
Microsatellites DNA markers

Blackfin tuna (Thunnus atlanticus)
Twenty homologous microsatellite markers, or simple sequence repeats (SSRs), are developed for the first time for blackfin tuna and tested on a sample of 35 specimens collected offshore La Martinique through the use of a direct Seq-to-SSR approach. The number of alleles per locus ranged between 5 and 26, and the expected heterozygosity ranged between 0.640 and 0.969. Three loci displayed significant departure from Hardy-Weinberg equilibrium expectations, likely reflecting occurrence of null alleles. Another locus showed consecutive alleles that differed by one base pair only. Consequently, this locus may be prone to elevated rates of scoring errors. More details in Antoni et al., 2014 Fisheries Bulletin 112, 322–325.
The remaining 15 loci are currently used to assess the population genetic structure and the degree of connectivity of the different stocks within the Atlantic basin species range. Adult blackfin tuna is a small tuna found in tropical and subtropical waters of the Western Atlantic Ocean. Over the last decade, the species has been increasingly exploited by fisheries in several parts of its range. Despite of this rising interest, management of the stock is still nonexistent.
As such, knowledge of stock structure is required to develop coordinated management of this resource accounting for subunits of the stock if they exist.
Adult black fin tuna from 7 locations including North Carolina, Gulf of Mexico, Florida Keys, La Martinique, Venezuela, Puerto Rico and Brazil, encompassing most of the species range have been sampled 2013 and 2014, although we have only completed data acquisition for 4 locations.
Preeliminary results show that pairwise FST estimates are overall very small, but some values are statistically significant, revealing for the first-time occurrence of significant spatial structuring of blackfin tuna populations and suggesting that management of the resource will require accounting for multiple stocks.
Isolation-by-distance is suggested by the outcome of the Mantel test (Fig.2) but the dataset does not allow excluding occurrence of barriers to gene flow (e.g. a role of the amazon barrier) and a more complex pattern of population structure. Further clarification of the status of U.S. populations particularly through increased sample sizes along the east coast (North Caroline site) and characterization of blackfin tunas from the Florida Keys are warranted and in progress. The finding of a possible isolation-by-distance model of population structure is unusual in tuna species and additional sampling in the Caribbean and South America would be useful to further examine this hypothesis and the potential role of larval pelagic dispersal.
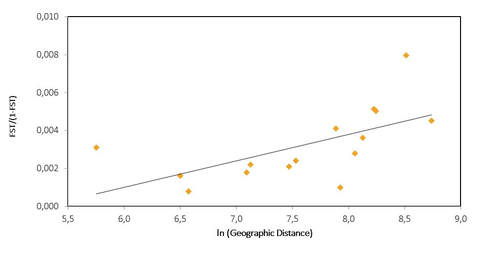
Plot of genetic (FST/(1-FST) versus ln (geographic distance) among 6 geographic populations of blackfin tuna.
Yellowfin tuna (Thunnus albacares)
In the Atlantic Ocean, multiple spawning locations have been reported for this warm water pelagic species, but a single stock is assumed during assessment efforts based on the observation of a continuous distribution of the species in tropical waters and the results of mark-recapture studies. Information on genetic stock structure is essential for management of this resource but is still very limited. In this work, 24 new microsatellite markers are developed and characterized on a sample of 35 yellowfin tuna specimens using Illumina paired-end sequencing and a direct ‘Seq-to-SSR’ approach. The number of alleles per locus ranged from 4 to 29 and estimates of expected heterozygosity were from 0.473 to 0.948 (n=35). Genotype frequencies at 8 of the microsatellites departed significantly from Hardy–Weinberg expectations, possibly reflecting occurrence of null alleles. More details in Antoni et al., 2014. Conservation Genetic Resources 6, 609–611.
The remaining 15 loci are selected based on their suitability for population genetic studies. Specimens are collected from 4 locations in the Western Atlantic (Northern Gulf of Mexico, U.S. Mid Atlantic and North Atlantic regions, and Venezuela) and two localities in the Eastern Atlantic (Ghana and Senegal). Sexually mature adults are targeted in priority during sampling in order to provide information on the breeding structure of the species. Samples are currently being genotyped and will be incorporated in an analysis of stock structure and connectivity in the Atlantic Ocean.
